鼻行动物几乎没有敌人。在岛屿内部,除了已经提到的沼泽鼩(Limnogaloides)之外,唯一的温血生物是Hypsiboas10属的鸟类(扩音器鸟)。所有这些都有鸣禽的大小,并且占据了与鼻行动物截然不同的栖息地。根据Bouffon和Sotprimarsch的说法,它们是海燕的后代,事实上是接近Hydrobates的物种。爬行动物缺乏。只有一种原始两栖动物物种(Urobombinator submersus),其巨大的幼虫在仪式宴会上被霍卡哈奇人吃掉。行动迟缓的Nasobema物种在Tyrannonasus12属的捕食性鼻行动物中有自己的敌人。然而,该属仅限于少数岛屿。在大多数情况下,只有在某些季节在一些小岛上繁殖的海洋鸟类偶尔会抓住鼻行兽。然而,生活在海岸上的那些物种(例如蜜尾兽和柱鼻兽)受到保护,免受鸟类的攻击,部分是通过毒液装置,部分是不可食用的;一般来说,鼻跃兽非常敏捷,不会被这些鸟抓住。
在这里,我们可以注意到哈伊艾伊动物群的另一个特点:昆虫表现出许多非常原始的形式。因此,蟑螂样种类由许多结构不同的类型代表,其中大多数可以归入蜚蠊科。除此之外,还有一些更高级的昆虫,尤其是膜翅目,而鳞翅目则完全不存在。因此,授粉部分是由膜翅目昆虫完成的(首先是由假炸弹属物种完成的,它们外表看起来像大黄蜂,但实际上与木蜂有关),部分是由石蛾和蟑螂完成的。没有蚂蚁。
作为一个显著的特点,应该提到六翼昆虫(六翼超目六翼类13);它们是古网翅目的后代,有陆生幼虫。这些昆虫大多是开阔地带的动物;也就是说,除了少数物种,它们避开了覆盖在较大岛屿山坡上的茂密原始森林。在这里,我们也可以记录较大岛屿有一些特有物种的特点。这些原始形态在较小的岛屿上完全缺失。这可能是由于较小的岛屿(例如Owmvussa或Sawabisi)是珊瑚岛,因此是最近形成的,或者是因为它们没有为笨拙的飞行者提供足够的防风保护,因此随着岛屿的沉没和变小,那里的特有物种灭绝了。
关于鼻行兽的系统布置,以下考虑因素适用:
如仍然用四条腿行走的单一物种(Archirhinos属=原始鼻行兽)所示,它们一定是从原始食虫动物进化而来的。在这方面,Limnogaloides在Mairuvili的存在意义重大;对于这种毫无疑问属于食虫动物的动物来说,它与Archirhinos有许多共同的特征,因此这两个物种可以追溯到共同的祖先并非不可能。
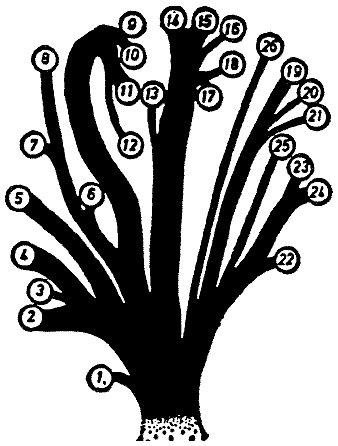
对于其余的,鼻行兽的系统分类主要遵循鼻子的发育程度。图2中的家谱显示了该目的系统细分,由Bromeante de Burlas(1950)提出。根据它,他区分了用腿行走的单鼻鼻行兽=Monorrhina pedestria (Archirhinus haeckelii是唯一的物种),用鼻行走的单鼻鼻行兽=Monorrhina nasestria (软鼻鼻行兽=Asclerorrhina和鼻足兽=Sclerorrhina),多鼻鼻行兽=Polyrrhina (短鼻鼻行兽=Brachyproata,长鼻鼻行兽=Dolichoprata)。尽管在这个方案中,大多数属都可以毫无困难地适应,但对于鼹鼠鼻行兽=Rhinotalpiformes,它们是否可以与穴居鼻行兽=Hypogeonasida14合并为一个群体,或者它们是否来源于Sclerhrohina(鼻足兽)并具有二次扩张的鼻器,目前尚不确定。
[图2]鼻行动物各属的拟议家谱(在Stultens数据的帮助下,从Bromeante de Burlas修改而来)。
1. Archirrhinos; 2. Nasolimaceus; 3. Emunctator; 4. Dulcicauda; 5. Columnifax; 6. Rhinotaenia; 7. Rhinosiphonia; 8. Rhinostentor; 9. Rhinotalpa; 10. Enterorrhinus; 11. Holorrhinus; 12. Remanonasus; 13. Phyllohoppla; 14. Hopsorrhinus; 15. Mercatorhinus; 16. Otopteryx; 17. Orchidiopsis; 18. Liliopsis; 19. Nasobema; 20. Stella; 21. Tyrannonasus; 22. Eledonopsis; 23 Hexanthus; 24. Cephalantus; 25. Mammontops; 26. Rhinochilopus.
树枝的粗细表明了几个属的相对物种数量。在这里,其他地方被视为独立属的Dulciauda和Dulcidauca被放在一起归入Dulciauda。