兰花鼻行兽(Orchidiopsidae)可以追溯到hopsorrhine的祖先,它们将陆地的生活方式换成了树上的生活方式,但已经拥有了各种各样的hopsorrhine式减少,特别是后肢的消失。因此,再也无法从它们身上培养出熟练的攀爬者。相反,一定是开始了一个进化过程,在这个过程中,动物不再从一个地方跳到另一个地方,而是在前肢和尾巴的帮助下慢慢爬行。无论如何,具有功能性鼻腿的代表性已经不为人所知,在它们的习性和构造上,今天的Orchidiopsis和Liliopsis物种是如此高度专业化,以至于最初甚至没有考虑它们由鼻跃兽进化而来(参见Gaukari Sudur、Bouffon和Paigniopoulos)。然而,与此同时,我们收集到了一些具有进化意义的发育数据,从中可以清楚地看出,没有矛盾的可能性,一个鼻
nasur和一个nasibia以及rhinanges最初是在Orchidiopsis的胚胎鼻中形成的,但后来被吸收了,因此完全发育的动物的鼻必须被视为是二次软化的。(Bouffon和Zapartegigarri 1953年写道: Les embryons des Orchidiopsides ne manquent point m de nasur ni de nasibie, mais pendant le develop fement [longueur de lembryon eniiron 15 a 18 mm] il y a lieu un ramollusement progressif de ceux-ci, de telle facon, que le nouveau-ne ne montre plus ancune trace dossification dans son nasarlum epiati et petaloide.*)
我们特别感谢Bouffon和他的学校对Orchidopsis组织特性的进一步解释。因此,Bouffon和Lo Ibilatze-Sudur能够证明Orchidiopsis物种的分泌引诱剂(粘液吸附剂)不是由外鼻表面形成的,事实上,外鼻表面缺乏任何可能参与其中的腺细胞。相反,它是由内表面的大腺体区域沿着鼻腔通道产生的,然后手动散布在鼻子的外表面。此外,同一作者成功地证明了Orchidiopsis 和Hopsorrhinus的尾巴抓握机制是同源结构。最后,Bouffons的学生Asteiides能够证明,在orchidiopsids的鼻子里有鼻足伸肌的痕迹,在胚胎发育过程中具有hopsorrhine类鼻行动物的位置特征。
*翻译:(Bouffon和Zapartegigarri 1953年写道;胚胎orchidiopsids既没有nasur,也没有nasibia.,但在发育过程中(胚胎长度为15至18毫米),这些经历了渐进的软化,使得新生动物在其扁平的花瓣状鼻中不再显示出任何骨化的痕迹。)
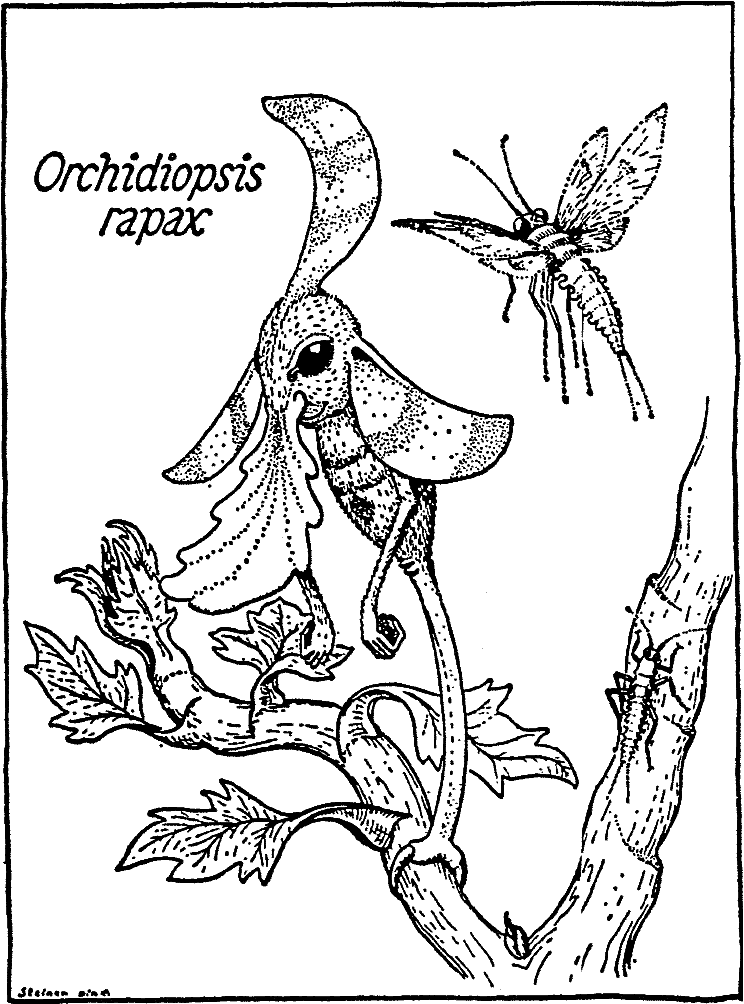
该科最著名的代表Ankers香草味兰花鼻行兽,Orchidiopsis rapax38生活在Mitadina岛的森林中,特别是在较高树冠的高度,但也在更温和的海拔地区,那里的风灾或洪水在树林中留下了开口(图九)。通常,这种动物一动不动地用尾巴站立,因此从远处看,它有点像一朵大花。这种相似性的起源是,巨大的耳朵、中部真皮冠和扁平的鼻子从头部突出,颜色鲜艳,像花瓣一样,而不显眼的绿色躯干起初并不明显。前面提到的鼻子上的引诱剂分泌物闻起来有香草味,起到了有气味的引诱作用。栖息在鼻子上或在鼻子上方扑动的昆虫会像闪电一样被细长手臂末端的可抓握爪子抓住,然后被送到嘴里。在前爪和有两只抓爪的尾巴的帮助下,位置偶尔会以变色龙般的缓慢变化。至于一个物种中几个个体之间的相互关系,我们一无所知;然而,到目前为止,已经采集了大约十几个携带不同发育阶段胚胎的怀孕标本(参见上文)。
nasur和一个nasibia以及rhinanges最初是在Orchidiopsis的胚胎鼻中形成的,但后来被吸收了,因此完全发育的动物的鼻必须被视为是二次软化的。(Bouffon和Zapartegigarri 1953年写道: Les embryons des Orchidiopsides ne manquent point m de nasur ni de nasibie, mais pendant le develop fement [longueur de lembryon eniiron 15 a 18 mm] il y a lieu un ramollusement progressif de ceux-ci, de telle facon, que le nouveau-ne ne montre plus ancune trace dossification dans son nasarlum epiati et petaloide.*)
我们特别感谢Bouffon和他的学校对Orchidopsis组织特性的进一步解释。因此,Bouffon和Lo Ibilatze-Sudur能够证明Orchidiopsis物种的分泌引诱剂(粘液吸附剂)不是由外鼻表面形成的,事实上,外鼻表面缺乏任何可能参与其中的腺细胞。相反,它是由内表面的大腺体区域沿着鼻腔通道产生的,然后手动散布在鼻子的外表面。此外,同一作者成功地证明了Orchidiopsis 和Hopsorrhinus的尾巴抓握机制是同源结构。最后,Bouffons的学生Asteiides能够证明,在orchidiopsids的鼻子里有鼻足伸肌的痕迹,在胚胎发育过程中具有hopsorrhine类鼻行动物的位置特征。
*翻译:(Bouffon和Zapartegigarri 1953年写道;胚胎orchidiopsids既没有nasur,也没有nasibia.,但在发育过程中(胚胎长度为15至18毫米),这些经历了渐进的软化,使得新生动物在其扁平的花瓣状鼻中不再显示出任何骨化的痕迹。)
该科最著名的代表Ankers香草味兰花鼻行兽,Orchidiopsis rapax38生活在Mitadina岛的森林中,特别是在较高树冠的高度,但也在更温和的海拔地区,那里的风灾或洪水在树林中留下了开口(图九)。通常,这种动物一动不动地用尾巴站立,因此从远处看,它有点像一朵大花。这种相似性的起源是,巨大的耳朵、中部真皮冠和扁平的鼻子从头部突出,颜色鲜艳,像花瓣一样,而不显眼的绿色躯干起初并不明显。前面提到的鼻子上的引诱剂分泌物闻起来有香草味,起到了有气味的引诱作用。栖息在鼻子上或在鼻子上方扑动的昆虫会像闪电一样被细长手臂末端的可抓握爪子抓住,然后被送到嘴里。在前爪和有两只抓爪的尾巴的帮助下,位置偶尔会以变色龙般的缓慢变化。至于一个物种中几个个体之间的相互关系,我们一无所知;然而,到目前为止,已经采集了大约十几个携带不同发育阶段胚胎的怀孕标本(参见上文)。











