亚族:Hypogeonasida (泥鼻行兽),
科:Rhinosiphonidae(广义虹吸鼻行兽),
属:Rhinotaenia (带鼻行兽),
2种
属:Rhinosiphonia(狭义虹吸鼻行兽),
3种
科:Rhinostentoridae(广义喇叭鼻行兽),
属:Rhinostentor(狭义喇叭鼻行兽),
3种
Hypogeonasida本身也是一个相对孤立的群体。它们都是不起眼的小动物,最初有一种地下生活方式,最典型的是Rhinotaenia属。接下来将描述作为特征代表的Rhinotaenia asymmetrica(浮潜带鼻行兽)。
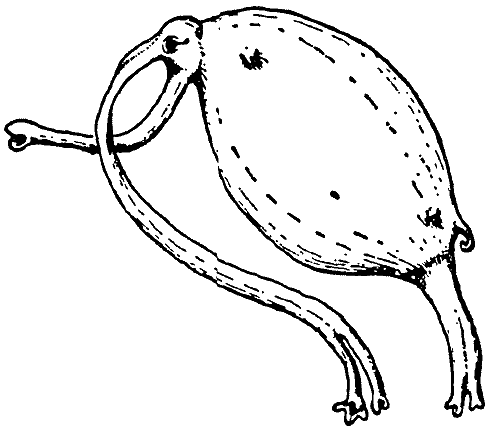
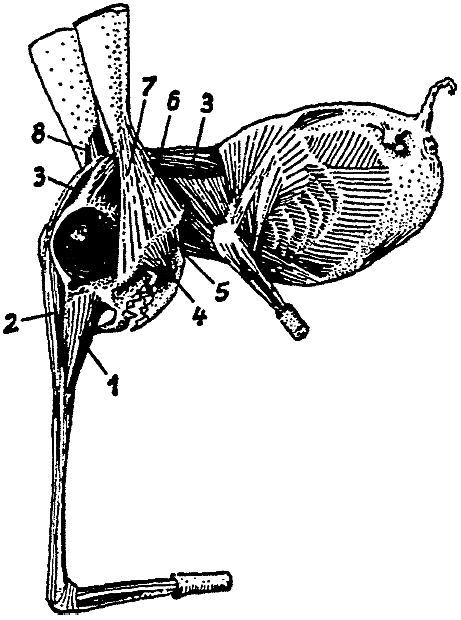
这种动物生活在几个小湖的泥地里,就像生活在几条缓缓流淌的溪流的近岸地区一样。在那里,它主要以寡毛类和幼虫昆虫为食,用长长的长鼻状嘴挖出并吞咽。在这样做的时候,Rhinotaenia每天都会向前挖掘一两码,深度约为一英尺。呼吸是通过长鼻子实现的,长鼻子像虹吸管一样延伸,长达16英寸,即长达动物头部和身体长度的四倍(见图3)。鼻子的不对称性——尽管气管很长,但左鼻孔及其末端玫瑰花结用于吸气,右鼻孔用于呼气,提供了完美的空气供应。
关于交配和繁殖,我们一无所知。全年都有怀孕的雌性和非常小的幼崽。
Beilig能够从分离的Rhinotaenia鼻子中提取出一种与Emuncator相同的粘蛋白。从形态学上讲,可以提出太多的观点,认为hypogeonasids起源于类似Emunctator的祖先(参见Bromeante de Burlas 1952以及Jerker和Celiazzini 1953)。
[图3]Rhinotaenia asymmetrica。(原图)

科:Rhinosiphonidae(广义虹吸鼻行兽),
属:Rhinotaenia (带鼻行兽),
2种
属:Rhinosiphonia(狭义虹吸鼻行兽),
3种
科:Rhinostentoridae(广义喇叭鼻行兽),
属:Rhinostentor(狭义喇叭鼻行兽),
3种
Hypogeonasida本身也是一个相对孤立的群体。它们都是不起眼的小动物,最初有一种地下生活方式,最典型的是Rhinotaenia属。接下来将描述作为特征代表的Rhinotaenia asymmetrica(浮潜带鼻行兽)。
这种动物生活在几个小湖的泥地里,就像生活在几条缓缓流淌的溪流的近岸地区一样。在那里,它主要以寡毛类和幼虫昆虫为食,用长长的长鼻状嘴挖出并吞咽。在这样做的时候,Rhinotaenia每天都会向前挖掘一两码,深度约为一英尺。呼吸是通过长鼻子实现的,长鼻子像虹吸管一样延伸,长达16英寸,即长达动物头部和身体长度的四倍(见图3)。鼻子的不对称性——尽管气管很长,但左鼻孔及其末端玫瑰花结用于吸气,右鼻孔用于呼气,提供了完美的空气供应。
关于交配和繁殖,我们一无所知。全年都有怀孕的雌性和非常小的幼崽。
Beilig能够从分离的Rhinotaenia鼻子中提取出一种与Emuncator相同的粘蛋白。从形态学上讲,可以提出太多的观点,认为hypogeonasids起源于类似Emunctator的祖先(参见Bromeante de Burlas 1952以及Jerker和Celiazzini 1953)。
[图3]Rhinotaenia asymmetrica。(原图)