树栖短吻鳄
(Ascendosuchinae)
鳄形目动物起源于陆生动物,其中一些在几个不同的场合变成了杂食性或植食性。在该群体的掠食性半水生成员中,在其进化史上多次趋同于完全陆地生活方式。
最终,一群小短吻鳄做了这两件事。
众所周知,现代鳄鱼偶尔会吃水果,而猫大小的变鳄则更进一步。随着盔甲的减少和更直立的姿势,这些短吻鳄是像狐狸一样快速奔跑的杂食动物,以各种各样的食物来源为食,植物物质在它们的饮食中占很大比例。
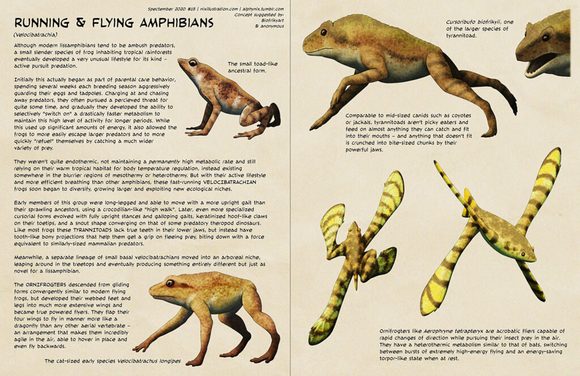
[图片:现代短吻鳄的小型陆生后代,有短而钝的鼻子和长而直立的四肢。]
Varigator cursorius
变鳄还保留了祖先偶尔爬上树的习惯,借此机会在树枝上晒太阳,避开更大的地面捕食者,或者从上面调查它们的领地。最终,这种行为导致了进化上的分歧,越来越多的树栖逃鳄从陆生形态中分裂出来。
为了越来越多地适应攀爬,逃鳄开发了可对握的抓握指、半抓握的尾巴,甚至更轻的稀疏装甲身体。它们在树冠上吃了越来越多的叶子、坚果和水果,它们的牙齿特化成一种令人惊讶的哺乳动物般的排列,口腔前部有锋利的牙齿,后部有多尖的咀嚼臼齿。
这些树栖鳄鱼只有老鼠那么大,行动缓慢,颜色神秘,很少见到。
[图片:一只变鳄在树枝上休息。]
树上的变鳄。
第2页
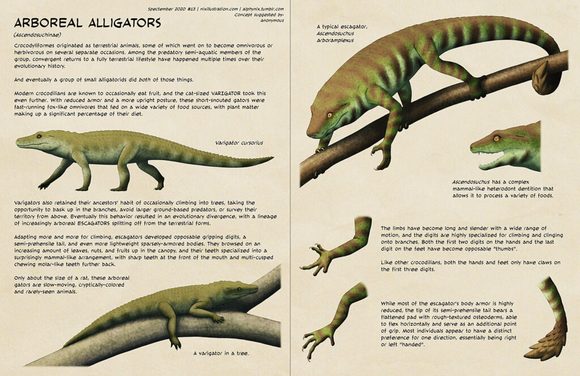
[图片:陆生短吻鳄的后代,紧紧抓住一根树枝。这是一种有点像变色龙的爬行动物,四肢细长,手和脚上有相对的手指,尾巴半可缠绕,盔甲减少。它的脸有一个短而宽的鼻子,眼睛朝前。]
一种典型的逃鳄,Ascendosuchus arboramplexus
[图片:逃鳄张着嘴的头部细节,显示出其哺乳动物般的分化牙齿。]
Ascendosuchus具有复杂的类哺乳动物异齿列,使其能够加工各种食物。
[图片:逃鳄四肢的细节,显示了手上的“双拇指”和脚上相对的第四个脚趾。]
四肢变得又长又细,运动范围很广,手指非常擅长攀爬和抓住树枝。手上的前两个指和脚上的最后一个趾都变成了可相对的“拇指”。
像其他鳄鱼一样,手和脚只在前三个手指上有爪子。
[图片:逃鳄尾巴尖端的细节,显示了覆盖着三角形鳞片状骨皮的宽垫。]
虽然大部分的逃鳄的装甲都大大减少了,但它的半可抓握尾巴的尖端有一个扁平的垫,上面有粗糙的骨皮,能够水平弯曲并作为额外的抓握点。大多数似乎对一个方向有明显的偏好,基本上是右撇子或左撇子。
(Ascendosuchinae)
鳄形目动物起源于陆生动物,其中一些在几个不同的场合变成了杂食性或植食性。在该群体的掠食性半水生成员中,在其进化史上多次趋同于完全陆地生活方式。
最终,一群小短吻鳄做了这两件事。
众所周知,现代鳄鱼偶尔会吃水果,而猫大小的变鳄则更进一步。随着盔甲的减少和更直立的姿势,这些短吻鳄是像狐狸一样快速奔跑的杂食动物,以各种各样的食物来源为食,植物物质在它们的饮食中占很大比例。
[图片:现代短吻鳄的小型陆生后代,有短而钝的鼻子和长而直立的四肢。]
Varigator cursorius
变鳄还保留了祖先偶尔爬上树的习惯,借此机会在树枝上晒太阳,避开更大的地面捕食者,或者从上面调查它们的领地。最终,这种行为导致了进化上的分歧,越来越多的树栖逃鳄从陆生形态中分裂出来。
为了越来越多地适应攀爬,逃鳄开发了可对握的抓握指、半抓握的尾巴,甚至更轻的稀疏装甲身体。它们在树冠上吃了越来越多的叶子、坚果和水果,它们的牙齿特化成一种令人惊讶的哺乳动物般的排列,口腔前部有锋利的牙齿,后部有多尖的咀嚼臼齿。
这些树栖鳄鱼只有老鼠那么大,行动缓慢,颜色神秘,很少见到。
[图片:一只变鳄在树枝上休息。]
树上的变鳄。
第2页
[图片:陆生短吻鳄的后代,紧紧抓住一根树枝。这是一种有点像变色龙的爬行动物,四肢细长,手和脚上有相对的手指,尾巴半可缠绕,盔甲减少。它的脸有一个短而宽的鼻子,眼睛朝前。]
一种典型的逃鳄,Ascendosuchus arboramplexus
[图片:逃鳄张着嘴的头部细节,显示出其哺乳动物般的分化牙齿。]
Ascendosuchus具有复杂的类哺乳动物异齿列,使其能够加工各种食物。
[图片:逃鳄四肢的细节,显示了手上的“双拇指”和脚上相对的第四个脚趾。]
四肢变得又长又细,运动范围很广,手指非常擅长攀爬和抓住树枝。手上的前两个指和脚上的最后一个趾都变成了可相对的“拇指”。
像其他鳄鱼一样,手和脚只在前三个手指上有爪子。
[图片:逃鳄尾巴尖端的细节,显示了覆盖着三角形鳞片状骨皮的宽垫。]
虽然大部分的逃鳄的装甲都大大减少了,但它的半可抓握尾巴的尖端有一个扁平的垫,上面有粗糙的骨皮,能够水平弯曲并作为额外的抓握点。大多数似乎对一个方向有明显的偏好,基本上是右撇子或左撇子。